 First the science. There are two principal lines of evidence to consider, the fossil record and modern molecular biochemistry. Charles Darwin expected the "tree of life" in the fossil record to look something like Fig. 1, with "species" constantly but gradually changing and occasionally splitting in two as branches appear and diverge. However, even he noticed a paucity of "transitional forms" in the fossil record. Instead fossilized species appear suddenly without obvious precursors or intermediate steps from the supposed previous branch of the tree, change very little over the millions of years and then fade out and (mostly) go extinct, with no obvious new species arising from them, as shown roughly in Fig. 2. Science articles often add horizontal dashed lines to such "trees" to show the supposed connections and transitions, but those do not represent the actual fossils.
First the science. There are two principal lines of evidence to consider, the fossil record and modern molecular biochemistry. Charles Darwin expected the "tree of life" in the fossil record to look something like Fig. 1, with "species" constantly but gradually changing and occasionally splitting in two as branches appear and diverge. However, even he noticed a paucity of "transitional forms" in the fossil record. Instead fossilized species appear suddenly without obvious precursors or intermediate steps from the supposed previous branch of the tree, change very little over the millions of years and then fade out and (mostly) go extinct, with no obvious new species arising from them, as shown roughly in Fig. 2. Science articles often add horizontal dashed lines to such "trees" to show the supposed connections and transitions, but those do not represent the actual fossils.The fossil record aspect most perplexing to Darwin was the so called Cambrian explosion. Some 540 million years ago (MYA), at the start of the Cambria
 n period of geological history, numerous new complex lifeforms, with sophisticated features like eyes, body plans, legs, swimming, mouths and guts, nervous systems, etc. burst into existence over just a few million years, with no apparent precursors from the previous Ediacaran age, which recorded only simple worms, algae mats, sponges, and the like as fossils. Darwin hoped that subsequent fossil finds would fill in the blanks, but that has not happened for the most part, and the same is true all through the fossil record.
n period of geological history, numerous new complex lifeforms, with sophisticated features like eyes, body plans, legs, swimming, mouths and guts, nervous systems, etc. burst into existence over just a few million years, with no apparent precursors from the previous Ediacaran age, which recorded only simple worms, algae mats, sponges, and the like as fossils. Darwin hoped that subsequent fossil finds would fill in the blanks, but that has not happened for the most part, and the same is true all through the fossil record.This conundrum for gradualist evolution remains as true today as it did 150 years ago, with no credible naturalistic explanation. Indeed, the problem was so clear in 1972 that Niles Eldredge and Stephen Jay Gould suggested the Punctuated Equilibrium (PE) model for evolution, claiming that the transitions happened so quickly that none of them were fossilized, and that the new species became populous as equilibrium forms, with little of no further phenotype (physical) changes until the next punctuation hit. Not only does this go against Darwin's clear requirement that there be no sudden major changes ("saltations" he called them), but it also goes against population genetics, which shows it is much harder to get major mutation changes - the kind needed to create a new species - over short period and in a small population, as required by the PE theory.
On the other hand, ID says, of course species remain largely fixed phenotypes as long as they exist, because the Darwinian mechanism, working in known ways on genomes, cannot create new features or the major genetic changes needed for totally new species. Many - perhaps most - species have one or more gene unique to them, with nothing similar in any other species. These so called orphan genes cannot be explained by a Darwinian mechanism (random mutation, plus natural selection) working on precursor genomes in any reasonable geological time. Any new feature in the fossil record would have required many new proteins (and hence genes) to construct the feature, and others to integrate it into the lifeform and make it functional, thereby rendering the transition even less likely for any purely natural process.
This finding of molecular biochemistry leads to the second piece of evidence for the process I will outline below. Studies of extant species demonstrate that for the most part, any traceable genetic effects leading to new species have arisen by devolution, that is, the damaging or removal of one or more genes in a pre-existing genome. Hence polar bears, for example, evolved from brown bear cousins by damaging the genes that limited their fat intake, allowing them to eat fatty seals without health problems, and by damaging the gene for melanin that produces dark pigments, leaving their fur white. Presumably such mutations first occurred in brown bears, but were not beneficial to them. However, when the polar bears forebears migrated into the arctic, those mutations were beneficial, helping the polar bear thrive in the new environment, and so were selected and then fixed into the growing population.
Yes, this is a clear case of Darwinian evolution producing a new species, just as Darwin claimed. Sometimes, Darwinism actually works! Note however, that although beneficial, those mutations damaged the genome, knocking out or damaging multiple genes. No new genetic information arose. Michael Behe, in his book Darwin Devolves describes this effect in depth. The same finding applies to almost every known case of beneficial mutation. Bacteria become resistant to antibiotics by damaging the genes that generate the cell-wall portals through which the antibiotics attack them. This is like an army burning its bridges behind them during a retreat: yes, it saves your army and slows down the enemy, but only by destroying part of your infrastructure that's needed in normal life.
Behe describes numerous examples, his first rule of adaptive evolution is to, "break or blunt any functional coded element whose loss would yield a net fitness gain". In hindsight, this effect should have been obvious. For any given gene, damaging mutations are far more likely than ones that might improve the gene somehow. Any new feature or function in an organism requires new proteins or enzymes, which therefore require new genetic information, and not just one or two minor changes. Most genes are hundreds of nucleotides long, and the probability of a new functional gene arising de-novo, by random mutations is all but negligible. On the other hand, a single point mutation in a gene is often enough to damage or render inoperative the resulting protein molecule. Thus, if a benefit to the species can be had by knocking out or damaging a particular gene, it is easy to do - just wait for almost any mutation to that gene - and very effective. It is easier and quicker to burn your bridges than to develop new weapons and rebuild your army to counterattack. Other research supports Behe's arguments and conclusions.
So Darwinism works, at least in this very limited sense. However, evolution by damaging the genome does not get us new biological features and functions. The polar bear is still a bear, and indeed could still mate with brown bears in principle. Devolution can only take you so far. Another example is dogs, which apparently evolved from wolves, via selective breeding - not quite natural selection, but the same rule applies. Most of the variations and new breeds of dogs have arisen by knocking out or damaging genes, or gene variants (alleles) found in wolves. Sure you get new breeds, but all dogs are still dogs - no new features - and one does not have to think too long about how most dog breeds would survive in the wild competing with wolves (natural selection). By damaging or removing parts of the genome, you can indeed get somewhat different life forms, but you cannot get anything completely new. For that, you need to add new genetic information, new genes that work together for new functions or phenotypic features. The Darwinian process of random mutations and natural selection simply cannot accomplish that.
In addition to the Cambrian explosion, there were other brief periods (almost moments) in geological time when whole batches of new life forms came into existence suddenly. Some examples include the Avalon explosion, and the bursts of new life forms following mass extinction events. Naturalist Darwinians claim these extinctions opened up numerous ecological niches which were then filled with newly evolved life forms. Yet they have no mechanism to credibly account for where the new genetic information came from for these new species that suddenly pop into existence.
The evolution model I would like to propose to account for the tree of life as it appears in the fossil record, builds on these findings of the last century or so. Figure 3 sketches what I have in mind, essentially an expansion of the PE model, with ID added in to account for the punctuations. Most of the time life goes on without much change. Species remain largely unchanged with some going extinct, and only occasional "new" species arising via devolution. These new species are very similar to their precursors. Think of all the dinosaurs, for example; numerous ones look very similar to each other, changed only in size or minor tweaks to their bones, yet each one gets a new name when unearthed. This is Darwinism at work, tweaking the world a little bit over millions of years, while the fossil record remains in equilibrium mode or "stasis".
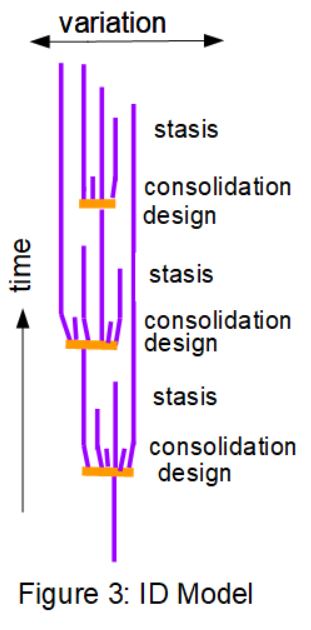 Then, at detectable moments in geological time, blasts of new species, genera, or even families arise almost suddenly, as "design" events, when new genetic information is injected into the biome. A brief period of "consolidation" would occur as the new information is integrated and accommodated by the new life forms. Some of those die out immediately as non-viable, or too few in number. Others adjust their phenotype or morphology as the new genes take effect, and settle out as new species.
Then, at detectable moments in geological time, blasts of new species, genera, or even families arise almost suddenly, as "design" events, when new genetic information is injected into the biome. A brief period of "consolidation" would occur as the new information is integrated and accommodated by the new life forms. Some of those die out immediately as non-viable, or too few in number. Others adjust their phenotype or morphology as the new genes take effect, and settle out as new species.Numerous plant and animal species have no known transitions from prior species; bladderworts and squids for example. Even some of the iconic transitional fossil sequences like horses and whales are not what they are presented to be in evolution textbooks. The steps (saltations) between the supposed intermediaries are huge from a genetic perspective, requiring many genetic additions and long fixation times.
In the case of whales, for example, only a few million years separates the supposed Pakicetus, land-based starting point from the earliest known, fully-formed aquatic whale. Such a transition would require an enormous amount of new genetic information in the form of many new genes. Meanwhile, population genetic models suggest that the time required for even a single mutation to be "fixed" in a population of say, 10,000 whale precursors with a ten year reproduction cycle, could exceed that time frame. It is beyond belief that many thousands of just right mutations, arising randomly, could occur in that time frame, resulting in a totally new family of whale species.
It is much more reasonable to suggest that, at such moments, an intelligent agent interceded somehow, to develop an array of new species and introduce them into Earth's biosphere. How that could be done, whether by purposely adding to or modifying DNA in various extant species, or by "creating" new species using similar genetic building blocks, with a few additions, cannot be known at present, although I have previously speculated on how this might be done. The injections of new information would likely occur in small populations, which are then released into the wild. With low numbers and short times during these consolidations, few if any fossils would be saved for us to dig up, and the fossil record would appear the way it does in Figure 2.
With a batch of new genetic information added into various genomes, the initial "explosion" of different lifeforms would be huge. Many of those would not be viable, perhaps, depending on the distribution of the new genes, and many new "lines" would doubtless go extinct even before they got started, either due to detrimental effects, or the exigencies of life - most individual lives are lost by random chance rather than from being slightly less fit. Nevertheless, enough new life forms would survive the first few generations, and would quickly settle down, via Darwinian selection, into separate, stable, viable species. By the time these became numerous enough to get one or more preserved in the fossil record, they would be a thriving species, now in equilibrium with its environment. Hence the punctuations in the fossil record, leading to steady state or "stasis".
This model suggests a range of potential research areas. The fossil record is an obvious starting place. Look for and catalogue all the sudden eruptions of new life forms without apparent precursors. How often do they occur? Are they all at once for all of life on Earth, as in the Permian extinction, or are they smaller in scale, limited to one area of the globe, or one set of lifeforms at the genus or family level? The fossil record is well established, even if it is still filling in slowly, so the data to do this must exist, even if no one has looked at it in this way.
On the biomolecular side, continue to research and collect the genetic changes that have caused known speciation events. In some cases, the "molecular clock" (such as it is) for random mutations, can be used to estimate when certain genes, similar among different species, were once the same gene, suggesting a branch point in life's tree, or at least in the subsequent existence of that gene. Similar approaches can be used to estimate when a given gene, perhaps one unique to a particular species, first came into being. For example, by looking at the variation in a single, uniquely human gene among extant human beings, the "age" of that gene - when it first arose - based on the accumulated variations, can be estimated.
It is conceivable that, as the fossil record is looked at more closely, and as the picture of genetic histories becomes clearer, the groups, dates and possibly locations may start to overlap, pointing to when and where the injections of new genetic info occurred. This is obviously a long term project. There will be lots of "noise" and gaps in the data, but if such a consilience or agreement between fossils and molecules can be found, it would be a major discovery.
The third piece of research would be to use comparative genetics to estimate how much genetic info was injected at these supposed design interventions by the intelligent agent. For example, what features and functions existed in possible partial precursor species, and what additional genetic information would be needed to account for the added or different features seen in the new species that arose? This could be done by carefully examining what genes are needed for similar features or functions seen in extant species. None of this would be precise or definitive, of course, but it could be useful and could hint at how the information was added.
For example, if some newly arisen species had (or its extant descendants have) a set of genes required for some new feature, and it can be shown that those genes could have come from two different families that cannot breed together, then clearly, one good explanation is that the agent took existing genes from two disparate species, and combined them to produce a novel feature in a new species. This would rule out the small genetic tweaks approach to developing a new line of lifeforms.
One last area of research for the proposed model would be to look closely at what blind Darwinian mechanisms can actually achieve over geological time scales. That is, at what level of variation and adaptation among species can random mutation plus natural selection alone - without intelligent direction, or added genomic information - produce actual genomic and morphological changes, possibly including new species? Several natural mutational and genomic change mechanisms are well known, and mathematical models exist to explore their realistic effects over time in a population of some size. The influence of environment and competition can be added to these models to see how much blind evolution might be possible. Actually, a lot of this work has already been done, including some empirical lab work, much of it by ID proponents seeking to determine where the line between natural evolution and ID must be in the hierarchy of life forms. See Michael Behe's The Edge of Evolution, for example. Indeed, initial considerations of this sort were key to the inception of ID theory, but additional research could provide a lot more understanding and insight.
Instead of trying to debunk Darwinism on the one hand, or completely dismissing Intelligent Design on the other, we should combine them into a better scientific paradigm for evolution. This reflects Hegel's philosophy of thesis (Darwinism), antithesis (ID) and synthesis (the combined model) as applied to evolution. As I understand the current situation, some form of ID theory is gaining credibility among scientists of various stripes, and in various places. Once ID is allowed to exist peaceably alongside the 150 year sole reign of Darwinism and its own neo-synthesis, then the above research can begin in earnest, exploring the model offered in Figure 3. It may be that the above suggestions will lead nowhere or to a mishmash of uncertainty, but even that would tell us something. Surely, however, even the possibility of finding out something about when and how the intelligent agent was at work, would be a huge addition to our scientific knowledge about the origin and history of life on Earth, and our relationship with the creator and the cosmos.
No comments:
Post a Comment